肌动蛋白体外自组织复合纤维结构的观察
激光生物学报 第13 卷第6 期 2004 年12 月
张 军1 ,2 , 王远亮1 , 何创龙1 , 程 超1 , CHEN Xin-yong3
(1. 重庆大学教育部生物力学及组织工程重点实验室,中国重庆400044 ; 2. 重庆医科大学生物学教研室,中国重庆400016; 3. Laboratory of Biophysics and Surface Analysis , University of Nottingham , Nottingham NG7 2RD , United Kingdom)
摘要
利用原子力显微镜(atomic force microscope , AFM) 和透射电子显微镜(transmission electron microscope , TEM) 技术,研究了低浓度肌动蛋白在体外简单热力学体系中,形成的自组织复合纤维结构。肌动蛋白在体外通过自组织过程能够聚合形成大尺度的、离散的、复杂的聚集纤维体系,分散的单根微丝较少;在微丝稳定剂鬼笔环肽干预下,肌动蛋白通过受调控的自装配过程,主要形成分散的单根微丝,以及少量由单根微丝组成的微丝束和纤维分支等简单微丝聚集结构。
关键词
肌动蛋白;自组织;复合纤维结构;原子力显微镜;透射电子显微镜
中图分类号: Q245 ;Q-334
文献标识码: A
文章编号: 1007-7146 (2004) 06-0401-05
Observation on the Complex Structure of Actin Filaments Forming in Self-organization in vitro
ZHANG Jun1 ,2 , WANG Yuan-liang1 , HE Chuang-long1 , CHENG Chao1 , Chen Xin-yong3
(1. The Key Lab of Biomechanics &Tissue Engineering , Ministry of Education , Chongqing University , Chongqing 400044 , China ;2. Biology Department of Chongqing University of Medicine Sciences , Chongqing 400016 , China : 3. Laboratory of Biophysics and Surface Analysis , University of Nottingham , Nottingham NG7 2RD , United Kingdom)
Abstract
The transmission electron microscope and the atomic force microscope - the novel tool for surface structure exploration ,are employed to examine the structure of actin filaments forming in self-organization in vitro. It is observed that the actin can be po-lymerized into discrete and complicated filament complexes of tree-like branch structure and random coil filaments cluster in large scale in the simple thermodynamic solution without the addition of F-actin stabilizing reagents phalloidin. However in the presence of phalloidin , the polymerized filaments are mainly the single F-actin in dispersive distribution , and the intricate filaments made of mi-crofilaments are seldom found under the same experimental conditions.
Key words
actin ; self-organization ; complex structure of filament ; atomic force microscope ; transmission electron microscope
肌动蛋白(Actin) 是真核细胞内最保守、含量最丰富的蛋白质之一,作为细胞骨架和结构的主要成分,具有极其重要的细胞生理功能[1 ,2 ] 。肌动蛋白一般以球形肌动蛋白(G-actin) 和纤维性肌动蛋白( F-actin , 即微丝) 两种形式存在。G-actin 的一级序列通常由375 个氨基酸残基组成,分子量约为43 KD ,溶液中,在Mg2 + , K+ , Na + ,及ATP 诱导下能自聚合形成高分子量、右手双螺旋结构的F-actin[3 ,4 ] 。在体内,F-actin 能够通过与一系列胞内蛋白相互作用,以适应不同生理条件下的功能需要[5 ] 。肌动蛋白在真核细胞中担负了多种重要的生理功能[6~15 ] ,是细胞内最重要的蛋白质之一,因而自从被发现以来,其结构及功能研究成为细胞生物学和结构生物学的重要内容。不仅多种晶体形式的G-actin 原子结构已获得阐明[16~18 ] ,而且F-actin 的原子结构模型也通过纤维X-射线衍射[19 ,20 ] 和电子显微镜成像而得到建立[21~23 ] 。
但是,纤维性肌动蛋白的复杂聚集结构体系却极少引起关注(尽管细胞内的微丝聚集体系—肌纤维和微绒毛的超微结构已得到阐明) 。因而,我们利用原子力显微镜(一种已广泛应用于肌动蛋白结构、功能及动力学研究的新型表面结构分析仪器[24 ] ) ,和透射电子显微镜,对肌动蛋白体外简单热力学体系中形成的自组织复合纤维结构进行了观察。
1 材料和方法
1.1 材料
G-actin 提取自牛骨骼肌,纯化按照Spudich 和Watt 等人的方法改良[25 ] 。分离的肌动蛋白达电泳单点纯,储存于4 ℃的G-缓冲液中备用;Na-ATP、DTT 购自Sigma ,Tris-base 、Tris-HCl 购自Promega , 其余试剂均为国产分析纯;CSPM-2000we 型原子力显微镜为中国科学院化学研究所本原纳米仪器有限公司生产,使用NANOPROBETM NP 型Si3N4 探针。H-600 透射电子显微镜为日立公司产品,使用300 目的铜网,以醋酸铀为负染色剂。
1.2 方法
1.2.1 溶液配制 G-缓冲液(2 mmolPL Tris-Cl , pH7. 5 , 0. 2 mmolPL CaCl2 , 0. 5 mmolPL DTT , 0. 2 mmolPLATP) 和F-缓冲液(5 mmolPL Tris-Cl , pH 7. 5 , 2 mmolPLMgCl2 , 100 mmolPL KCl , 1 mmolPL DTT , 1 mmolPLATP) 均按常规方法制备,所有使用的溶剂和新鲜配制的溶液均通过直径0. 22 μm 的滤膜过滤去除颗粒。
1.2.2 原子力显微镜样品制备 用F-缓冲液将1ml 纯化G-actin 精确稀释至100 ml ,使溶液中G-actin的最终浓度为5μgPml ,然后将样品置于37 ℃恒温箱中孵育30 min 使其聚合。聚合完成后,在新鲜剥离的、经F-缓冲液浸润的云母表面(1 cm ×1 cm) 滴加5 μl 待测溶液,使蛋白能够良好分散。沉降15min 后,自然脱水以尽可能维持肌动蛋白真实的聚合结构,制成待测样品。
1.2.3 原子力显微镜成像 样品制备完成后不经过任何物理和化学处理,立即在室温下(25℃) 用原子力显微镜的接触模式成像。原子力显微镜采集的图像保存为BMP 格式,储存在电脑中做进一步的分析。上述所有操作均在超净工作条件下进行,以避免污染。
1.2.4 负染色样品的制备 实验组使用普通F-缓冲液,对照组加入过量鬼笔环肽到F-缓冲液中,使最终溶液中的鬼笔环肽与球形肌动蛋白的分子比达到5∶1 或以上,使聚合体系中的鬼笔环肽充分过量;分别用实验组和对照组的F-缓冲液将1 ml 纯化G-actin 精确稀释至100 ml ,使溶液中G-actin 的最终浓度为5μgPml ,然后将样品置于37 ℃恒温箱中孵育30 min 使其聚合。聚合完成后,在300 目的铜网上滴加适量待测溶液。沉降15 min 后,小心吸去多余的液滴,然后按照负染的常规方法制成待测样品。
1.2.5 透射电子显微镜观察 样品制备完成后,立即按照常规操作在H-600 (Hitachi) 透射电子显微镜上对样品进行观察和摄影。
2 结果
2.1 自组织复合纤维结构的原子力显微镜观察
肌动蛋白在体外通过自组织能够聚合形成大尺度的树状分支纤维丛结构和聚集的、相互缠绕的无规卷曲纤维簇。目前初步判断,观察到的聚集结构可能是由微丝通过非共价作用(氢键,范德华力,静电作用等等) 进一步聚合形成的有序结构;推测微丝间的相互缠绕或形成(多级) 螺旋可能是聚集纤维复合体的主要构成方式。
2.1.1 状分支结构的纤维丛 具有分形结构特征的树状分支结构,由多级纤维分支形成( Fig.1)。 暗示聚集的纤维结构可能是通过微丝之间相互作用而形成的多级螺旋的纤维复合体系。推测次级纤维成份之间的相互作用可能在树状分支结构的产生过程中,具有重要的作用,但复合纤维结构产生的详细聚合动力学机制还有待于进一步的深入研究。

图1. 树状分支结构
Fig.1 Tree-like branch structure
2.1.2 无规卷曲的纤维簇 在大尺度的范围内,还能观察到大量的纤维互相缠绕、簇集在一起,呈无规律的卷曲分布状况(Fig. 2) 。无规卷曲的纤维簇可能与树状分支结构的纤维丛有很强的同源性。二者可能是微丝进一步聚合过程中、不同聚集阶段的产物,具有不同的空间拓扑构象。
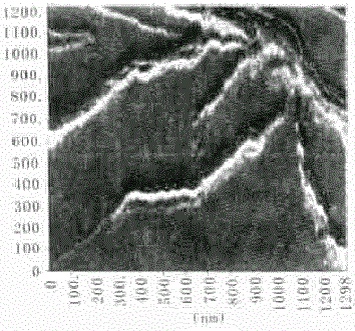
图2.无规卷曲的纤维簇图像
Fig.2 Random coil filaments cluster. 1
2.1.3 自组织纤维结构分支角度的频数分析 利用SPSS10. 0 对自组织纤维结构分支角度的频数分布进行了分析。发现,自组织纤维结构分支角度主要分布在35~65 度之间,在这个分布范围之外的纤维数量很少(Fig. 3) 。平均分支角度为52. 08 ±9. 42度,分支角度的大小反映了微丝之间相互作用的能级状态,集中分布的角度范围应该是微丝间相互作用的最低能量状态区域。
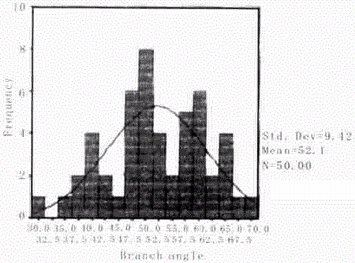
图3. 分支角度的频数分布图
Fig.3 Frequency distribution of branch angle
2. 2 复合纤维结构负染色的透射电子显微镜观察
在过量微丝稳定剂鬼笔环肽介入下的肌动蛋白体外聚合实验中,单根微丝为主要的聚合纤维成分,普遍较长,单独存在或在沉降过程中形成由取向不同的微丝组成的交联结构,还能观察到少量由数根微丝构成的微丝束和纤维分支等简单聚集结构,但观察不到大尺度的复杂纤维结构( Fig. 4a) ;肌动蛋白体外通过自组织过程主要形成离散的、大尺度的复杂纤维结分散的单根微丝很少。具有较粗直径的长纤维很常见,而且较长的纤维一般直径较粗,多有分支产生(Fig. 4 b , c) 。肌动蛋白自组织纤维结构负染色的透射电镜观察结果与原子力显微镜的结果完全一致。

图4. 肌动蛋白复合纤维透射电镜图
Fig.4 Images of negatively stained polymerized actin filaments in the presence of phalloidin and in self-organization.
(a) Image of actin filaments polymerized in the presence of phalloidin ( ×80000) ; (b) , (c) ones in self-organization ( ×80000) . The scale bar represents 100 nm.
3 讨论
肌动蛋白在F-缓冲液中,没有微丝稳定剂存在的条件下,能够通过自组织过程形成大尺度的、离散的复杂纤维聚集结构;而在常规性微丝工具药物(稳定剂) 鬼笔环肽介入下,肌动蛋白在F-缓冲液中,通过自聚合过程主要形成分散的单根微丝和少量由单根微丝组成的微丝束或纤维分支等简单微丝聚集体,不能形成复杂的纤维复合结构。推测,鬼笔环肽与微丝结合后,会占据肌动蛋白空间构象的特定位点,降低单根微丝的柔韧性[20 ] ,这可能会干扰或抑制微丝间的进一步相互作用;而且在鬼笔环肽存在时,聚合形成的微丝普遍较长,微丝之间通过相互作用产生稳定的结构需要更大的空间位移,使空间位阻增大,进一步地阻止了复杂纤维聚集复合体的形成。在自组织过程中,肌动蛋白聚合形成的具有天然构象和适度长度的微丝易于通过相互作用,形成簇集的、大尺度的复杂纤维聚集结构。细胞内,微丝聚集结构———肌纤维和微绒毛均由平行的束状微丝构成,构成方式相对简单。在体外,肌动蛋白通过自组织过程形成的聚集纤维结构复杂得多,形态与细胞内的聚集结构有明显的差异,而且其构成方式还未能揭示,有待于进一步的深入研究。
肌动蛋白自组织动力学及其复合纤维结构反映了由一级结构所确定的肌动蛋白内在热力学特性。因而,对肌动蛋白在体外简单热力学体系中的自组织热力学和分子动力学进行深入的研究和探索,无疑能够为阐明细胞内与肌动蛋白功能密切相关的聚合动力学调控机制提供重要的理论依据和指导,并有助于理解生物大分子动力学、结构多态性和功能多样性之间的内在联系。
Reference
[1]POLLARD T D , COOPER J A. Actin and actin-binding pro-teins. A critical evaluation of mechanisms and functions[J ] .
Annu Rev Biochem , 1986 , 55 :98721035.
[2]COLUCCIO L M, BRETSCHER A. Reassociation of mi-crovillar core proteins : making a microvillar core in vitro[J ] .J Cell Biol , 1989 , 108 :4952502.
[3]ESTES J , SELDEN L , KINOSIAN H , et al. Tightly bounddivalent cations of actin[J ] . J Muscle Res Cell Motil , 1992 ,13 :2722284.
[4]SHETERLINE P , CLAYTON J , SPARROWJ C. Actin. In Protein Profile[M] . New York :Academic Press , 1995.
[5]FURUKAWA R , FECHHEIMER M. The structure , function and assembly of actin filament bundles[J ] . Internat Rev Cy-tol , 1997 , 175 :29290.
[6]HOLMES KC. A molecular model for muscle contraction[J ] .Acta Crystallogr A , 1998 , 54 :7892797.
[7]POLLAR T D. Actin[J ] . Curr Opin Cell Biol , 1990 , 2 :33240.
[8]SMITH SJ . Neuronal cytomechanics : the actin2based motility of growth cones[J ] . Science , 1988 , 242 :7082715.
[9]POLLARD T D , AEBI U , COOPER J A , et al. Actin structure , polymerization , and gelation [J ] . Cold Spring Harbor Symp Quant Biol , 1982 , 46 : 5132524.
[10]HELDMAN A W, GOLDSCHMIDT2CLERMONT P J . Cell signalling and motile activity [ J ] . Symp Soc Exp Biol ,1993 , 47 :3172324.
[11]GAVIN R H. Microtubule2microfilament synergy in the cytoskeleton[J ] . Int Rev Cytol , 1997 , 173 :2072242.
[12]ATENCIA R , ASUMENDI A , GARCIA2SANZ. Role of cytoskeleton in apoptosis [J ] . Vitam Horm , 2000 , 58 : 2672297.
[13]TITU M A , GILBERT S P. The diversity of molecular motors : an overview[J ] . Cell Mol Life Sci , 1999 , 56 : 1812 183.
[14]LUO L. Actin cytoskeleton regulation in neuronal morphogenesis and structural plasticity[J ] . Annu Rev Cell Dev Biol , 2002 , 18 :6012635.
[15]CRITCHLEYD R , HOLTM R , BARRY S T, et al. Integrin2mediated cell adhesion : the cytoskeletal connection [J ] . Biochem Soc Symp , 1999 , 65 :79299.
[16]KABSCH W, MANNHERZ H G, SUCK D ,et al. Atomic structure of the actin :DNase I complex[J ] . Nature , 1990 ,347 :37244.
[17]MCLAUGHLIN P J , GOOCH J T, MANNHERZ H G, etal. Structure of gelsolin segment 12actin complex and the mechanism of filament severing [J ] . Nature , 1993 , 364 :6852692.
[18]SCHUTT C E , MYSLIKJ C , ROZYCKI M D , et al. The structure of crystalline profilin2beta2actin [ J ] . Nature ,1993 , 365 :8102816.
[19]HOLMES K C , POPP D , GEBHARD W, et al. Atomic model of the actin filament [J ] . Nature , 1990 , 347 :44249.
[20]LORENZM, POPP D , HOLMES KC. Refinement of the F-actin model against x2ray fiber diffraction data by the use of a directed mutation algorithm[J ] . J Mol Biol , 1993 , 234 :8262836.
[21]BREMER A , MILONIG R C , SUTTERLIN R , et al. The structural basis for the intrinsic disorder of actin filament : the lateral slipping model [J ] . J Cell Biol , 1991 , 115 :6892703.
[22]BREMER A , HENN C , GOLDIE K N , et al. Towards atomic interpretation of F2actin filament three dimensional reconstructions[J ] . J Mol Biol , 1994 , 242 :6832700.
[23]MILLIGAN R A , WHITTAKE M, SAFER D. Molecular structure of F2actin and location of surface binding sites[J ] .Nature , 1990 , 348 :2172221.
[24]ZHANGJ , WANG YL , GU L , et al. Atomic force microscopy of actin [ J ] . Acta Biochim Biophys Sin , 2003 , 35(6) : 4892494 (in Chinese) .
[25]SPUDICH J A , WATT S. The regulation of rabbit skeletal muscle contraction. Ⅰ. Biochemical studies of the interaction of the tropomyosin2troponin complex with actin and the proteolytic fragments of myosin [ J ] . J Biol Chem , 1971 ,246 : 486624871.
作者简介
张军: 男,1973 年10 月出生于四川南充。博士,从事细胞生物学,稳恒磁场生物学效应和抗肿瘤药物细胞毒性等领域的研究工作。近5 年来已发表学术论文9 篇,被SCI 收2 篇,CA , CSCD 收录7 篇。Tel : 65102508(O) ; 65104805(H) ; Fax : (023) 65316247 ;E2mail : zhangjun1017 @sohu. com
Biography
ZHANG Jun : male , born in October 1973 , Nanchong , Sichuan province. He is a Ph. D. , majoring in cell biology , biological effects of static magnetic fields and cytological effect of anti2tumor reagents. He published 9 papers in the recent 5 years , two of which was cited by SCI , and seven by CA , CSCD.